Brain Size and Intelligence
Not too long ago, I was reading about the differences between the human brain and the brains of chimpanzees. It got me thinking about how we as humans somehow possess a greater level of intelligence than primates (at least, that we’ve observed) despite similar brain morphologies.
Mainly to satisfy my own curiosity, I wrote a piece about brain size and intelligence in humans versus other primates. It’s been written in a bit of a pop science style (thought it was worth giving it a try).
Enjoy!
To what extent is higher-level intelligence in humans (relative to primates) attributable to brain size (cm3)?
Outline
- Introduction
- Background Information
- Discussion
- Immediate implications of a larger brain
- White matter and grey matter
- The cranial radiator hypothesis
- Neurotransmitters and GLUD2
- Conclusion
Introduction
Human civilisation can be traced back through millennia, from the time of the Mesopotamians and Aztecs, to the world as we know it today. As humans, we have formed cultures and traditions that have survived for centuries. We have created areas of knowledge, like science and literature, for which we have designed frameworks and sub-domains for people to produce more knowledge. We build skyscrapers, each taller than the last, we ponder over investments, driven to earn, and we perform experiments, always eager to expand our understanding of the world around us.
Our ability to do all of these things has been well established, but why is it that the other animals that belong to the primate order cannot perform the advanced cognitive functions that underwrite those processes?
The answer is simple and somewhat intuitive. Of the discovered species of animals on the planet, we display the greatest capacity to process and integrate large amounts of incoming sensory information, and compare this new information with the existing information that we hold. We also can think conceptually, as well as in abstract terms. In short, we display higher-level intelligence.
So how did this unique intelligence that humans possess come about? Why is it that we can feel that almost electrical tension in the denouement of Macbeth, and an orang-utan cannot?
This essay seeks to evaluate the extent to which higher-level intelligence in humans (relative to primates) can be attributed to brain size by considering the primate order and looking at how human brains in particular have evolved.
Background Information
First, let us establish some facts about the primate order. As we can see from the cladogram below, humans and chimpanzees both diverged from gorillas before splitting into two separate species. Interestingly, out of all the great apes, humans are outliers in terms of chromosome number; we have 23 pairs of chromosomes whereas the rest of the great apes have 24 pairs. Our second chromosome was formed millions of years ago from the end-to-end fusion of 2 ancestral chromosomes (Ridley, 2000)
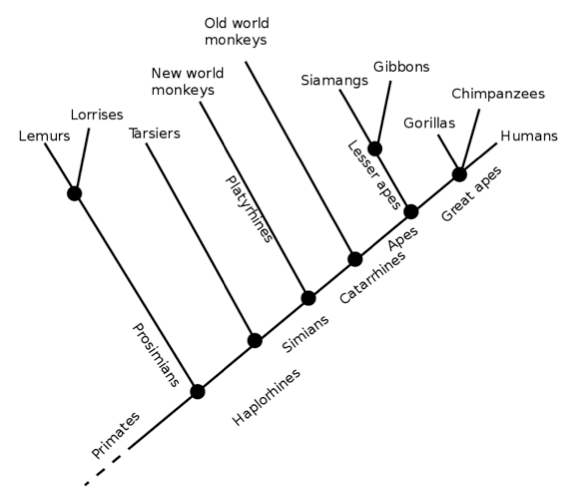
Following this divergence from the great apes (through the fusion of the 2 ancestral chromosomes), the animals in the homo genus (which initially started as australopithecines) underwent periods of evolutionary change, which gave rise to several distinct features, the most prominent of which is encephalisation – an increase in the volume of the brain. During the evolution of Homo sapiens, it is estimated that our brains were adding 150 million new neurons to its own vast array of neural circuitry every 100,000 years (Matt Ridley jovially labels the scientist who calculated this figure “a mathematical masochist”).
To translate this from the cellular to the physiological, the volume of the human brain increased nearly threefold, to its present volume of approximately 1400cm3 (Hofman, 2014). At the same time, the volume of grey matter and white matter in the brain increased, in conjunction with further cortical folding (the folding of the cerebral hemispheres). The brains of the great apes and primates, on the other hand, tend to be far smaller (350-450cm3), but display similar patterns of organisation in the brain; they too have the forebrain, which holds the cerebrum, the midbrain, in which the limbic system resides, and the hindbrain, where the medulla oblongata and cerebellum are found (Hofman, 2014).
Discussion
Immediate implications of a larger brain
Let us now move on to the question at hand, and consider the extent to which we can attribute the intelligence of modern day humans to brain size. As mentioned previously, it is clear that human brains are larger than the brains of other primates. So what does this higher brain size translate into?
The most obvious answer to this question is that a larger brain would have a higher number of functional neurons, as substantiated by the fact that humans have 86 billion neurons, whereas chimpanzees have 26 billion (Hofman, 2014). In particular, humans have a larger cerebral cortex, the centre for higher order processing in the brains of primates. The immediate result of a larger cerebral cortex is a greater number of neurons and nuclei dedicated to specific tasks, thus increasing the potential of the human brain to carry out a greater number of cognitive functions than our primate cousins.
White Matter and Grey Matter
However, it may not be the size of the cerebral cortex that has given the human brain an intellectual advantage over other hominids and primates. 50% of the human cerebral cortex (by volume) is composed of grey matter (Hofman, 2014). Grey matter consists of sections of the central nervous system that carry the cell bodies of neurons, their axons, their dendrites, the capillaries that nourish them, and most importantly, their synapses. The synapses are the points of communication between individual neurons. They are fluid filled gaps over which neurons will exchange chemical information to determine the direction over which an impulse will be propagated. In essence, they are connection points between different neurons, which may be associated with different nuclei (clusters of neurons with similar functions) in the brain, enabling different impulses from different parts from the brain to be linked and integrated to be sent to other nuclei. This forms the basis for higher order thinking – it is based on the summative effect of multiple pre-synaptic neurons producing a response in a particular postsynaptic neuron. In plain English, it allows us to synthesise new knowledge from multiple pieces of existing knowledge.
To sum up, the higher proportion of the human brain (relative to other primates) devoted to grey matter supports the argument that the size of the human brain may not be the main factor that has led to human intelligence.
However, we have to consider the fact that the human brain is, inexorably, larger than the brains of other primates. As a result, the higher proportion of the cerebral cortex dedicated to grey matter physically manifests as a higher volume of grey matter and therefore, a greater number of synaptic junctions, which forms more complex circuitry in humans than in primates. Hence, we have to consider the proposition that the higher-level intelligence that humans possess can be indirectly attributed to brain size.
Another point in favour of the latter argument relates to the proportion of white matter in the human brain. White matter, unlike grey matter, is comprised of the elongated axons that connect different nuclei. Hence, we can think of grey matter as holding the switches in the brain with the white matter forming the “wires” that connect a series of “switches” together.
As we can see below, the proportion of white matter as a function of brain volume in primates increases at a near exponential rate with brain volume.
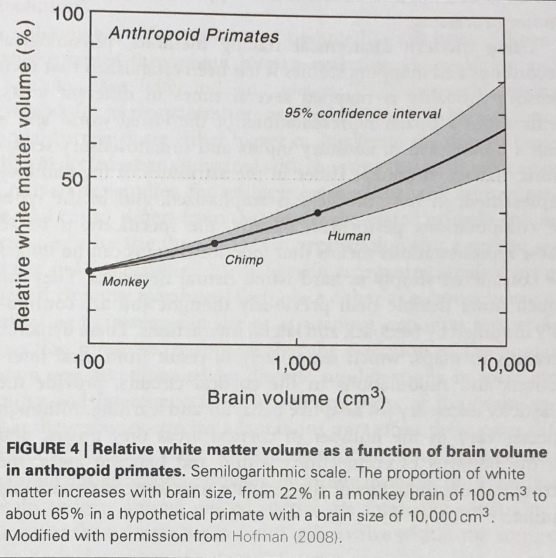
Does this correlation imply a causative influence conferred by brain volume? To answer this question, we must once again venture into the history of the evolution of the human brain.
As we know, there was a definite increase in the volume of the human brain as we diverged from primates. With this larger volume came with it a diaspora of different nuclei distributed around the vast neural landscape of the brain. This created a need to generate high interconnectivity between different nuclei, with short conduction delays. To strike a compromise between these two factors, the human brain evolved to create greater interconnectivity between nuclei by dedicating more of the brain to white matter, and reduced conduction delays by developing myelinated axons in the white matter.
Due to the development of this system, humans are able to integrate information from multiple sources seamlessly, establish patterns, and generate new information rapidly. We can therefore attribute these characteristics, which form part of what we define as “human intelligence,” to evolutionary changes that were necessitated by an increase in brain volume.
The cranial radiator hypothesis
On the other hand, brain size may not necessarily be the main reason why humans are the most intelligent primates. The human brain can only work as hard as it does, firing off countless action potentials in a second, because heat, a metabolic waste product of its activities, can be removed at such a rate that it does not inhibit brain function.
Unlike most other primates, humans have a shunt of veins that flow close to the surface of the cerebral cortex, helping to remove excess heat generated by the activity of the 86 billion neurons in the brain (Falk, 1990). This cranial radiator removes temperature as a limiting factor and facilitates greater neuron activity per unit time, thus increasing processing speed and possibly contributing to the development of human intelligence.
Neurotransmitters and GLUD2
Let us consider something we have touched on briefly, the neurochemistry of the brain, specifically the neurotransmitters found in primates. How do the neurotransmitters used in the human brain differ from those found in other primates? The minor role the size of the human brain plays in the development of human intelligence is most clearly illustrated when one considers the neurochemistry of the human brain. Although we share a large number of our neurotransmitters with other primates, we have different mechanisms to recycle them for later use. Could this be main reason why humans have higher-level intelligence than primates?
Take for example, the GLUD2 gene. The human version of the gene differs from that of other primates by simply 2 amino acids, but this change is sufficient in producing a more efficient version of the glutamate dehydrogenase (GDH) enzyme that GLUD codes for (Bradbury, 2005). GHD can catalyse the decomposition of glutamate (a neurotransmitter) into alpha ketoglutarate, or catalyse the reverse reaction. Thus, the more efficient GDH produced by the human GLUD2 gene gives rise to greater glutamate turnover, which could have facilitated a greater rate of synaptic transmission in humans relative to other primates.
Conclusion
Let us circle back to the question posed at the very beginning of this essay – to what extent can human intelligence relative to primates be attributed to brain size? Weighing all of the factors we have considered, it becomes clear that our larger brain size is, inexorably, the main reason we have a greater capacity to interpret and analyse information relative to primates. There is also an ineluctable correlation between brain size (cm3) and the proportion of white matter as well as grey matter in the brain. As a result, the larger size of the human brain allows for greater interconnectivity (brought about by a larger amount of white matter) between greater numbers of nuclei (conferred by a larger amount of grey matter).
Although the cranial radiator hypothesis accounts for higher neuronal activity, such activity would likely be impossible without the vast number of neurons already present in the brain, which exist in such high quantities due to the large volume of the brain.
However, the role of genes in giving rise to human intelligence through influencing the behaviour of neurons cannot be understated. Up to 100 genes are expressed differently in the human brain than in the brain of a chimpanzee. This piece of evidence lends itself to the idea that human intelligence could be a product of genetic factors, rather than morphological features. On the other hand, the two may not be mutually exclusive, as these genetic factors could also be responsible for the morphological differences in the brains of humans and other primates.
References + Further Reading
- Hofman, M.A., 2014. Evolution of the human brain: when bigger is better. Frontiers in neuroanatomy, 8, p.15.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3973910/
- Falk, D., 1990. Brain evolution in Homo: The “radiator” theory. Behavioral and Brain Sciences, 13(2), pp.333-344.
- Ridley, 2000. Genome. Howes.
- Bradbury, J., 2005. Molecular insights into human brain evolution. PLoS Biology, 3(3), p.e50.
https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.0030050